

Si la resistencia vascular es proporcional a la diferencia de presión entre una arteria y una vena, e inversamente proporcional al flujo en esa región, entonces la presión arterial (Pa) debería ser igual a la presión venosa (Pv) cuando el flujo hacia la región (Qin) es cero. Sin embargo, se ha demostrado repetidamente que Pa es mayor que Pv cuando Qin=0. (1-7) La posible importancia de esto para la medición de la resistencia vascular recibió una mayor atención después del análisis de Bellamy sobre las relaciones presión-flujo diastólicas en el corazón canino, en el que encontró que la presión cuando Qin=0 (es decir, la intersección de flujo cero en la relación presión-flujo, que se denominará Pz=0) era de 30-50 mmHg, mientras que la Pv solo era de 5-10 mmHg. Propuso que Pz=0 se debía a una cascada vascular (Vascular Waterfall) o al mecanismo de resistencia de Starling, tal como lo describieron Permutt y Riley a nivel de las arteriolas. En coherencia con esta hipótesis, factores como un aumento en la frecuencia cardíaca o intervenciones farmacológicas que incrementan el flujo coronario y deberían producir dilatación arteriolar, disminuyeron Pz=0. (Bellamy, 1978) Por lo tanto, razonó que el flujo coronario podría regularse mediante cambios en el diámetro de los vasos, es decir, cambios en la resistencia o en el tono arteriolar que causan variaciones en Pz=0. Además, según esta hipótesis, en un órgano con buena autorregulación, como el corazón, Pz=0 solo podría medirse en estudios dinámicos, ya que el tono cambiaría con disminuciones de presión en estado estable y Pz=0 se subestimaría. En apoyo a sus observaciones, se han encontrado valores de Pz=0 mucho mayores que Pv en estudios dinámicos de presión-flujo en el cerebro, (Dewey, 1974; Early, 1974) el riñón, (Ehrlich, 1984) y la extremidad amputada. (Ehrlich, 1980; Braakman, 1983).
Pronto, Spaan y Eng identificaron una posible falacia en este razonamiento, proponiendo que Pz=0 en estudios dinámicos se debe a la descarga de la capacitancia vascular en o después del punto de medición del flujo y la presión. Así, la resistencia aún debería calcularse a partir de la diferencia entre Pa y Pv, pero solo en condiciones de estado estacionario para evitar transitorios provocados por el flujo desde regiones elásticas a medida que la presión cae. Se intentó eliminar el flujo de capacitancia mediante diversas técnicas, (Dole 1982; Dole 1984; Kirkeeide, 1981; Aversano, 1984; Canty, 1987) como el análisis de presiones decrecientes y crecientes (Aversano, 1984) y el estudio de impedancia. (Canty, 1987; Sato, 1971) Sin embargo, estas técnicas no descartan cambios en la resistencia dependientes de la presión, que también pueden ser importantes. (Dole, 1984) Además, el hecho de que el flujo venoso continúe cuando el flujo de entrada es cero (Baez, 1974) no explica la capacitancia, ya que este flujo podría ser distal al resistor de Starling. Dada la importancia de la medición de la resistencia en estudios hemodinámicos y las implicaciones de las explicaciones alternativas de Pz=0 (resistencia de Starling versus efecto de capacitancia), esta controversia debe resolverse. Por lo tanto, se examinaron las relaciones presión-flujo arterial dinámicas en extremidades amputadas de perros aisladas in situ, en las cuales previamente se había encontrado un Pz=0 dinámico superior a 60 mmHg. (Baez, 1977)
Se aplicaron tres pruebas. Primero, la variación de Pv, donde se elevó Pv en pasos para determinar si los cambios en Pv por debajo de Pz=0 afectarían Qin o Pa. Si Pz=0 se debía a un mecanismo de resistencia de Starling o a una presión crítica, entonces elevar Pv no debería tener efecto hasta superar la presión crítica. Segundo, la variación del tiempo hasta flujo cero, donde se varió el tiempo para alcanzar flujo cero entre 1 y 10 segundos. Si Pz=0 se debía al tono arteriolar y a un mecanismo de resistencia de Starling, entonces Pz=0 no debería cambiar en tiempos cortos, ya que se cree que los ajustes autorregulatorios del tono arterial tardan aproximadamente 4 segundos en ocurrir. (Ehrlich, 1984; Dole 1982; Mosher, 1964) En tiempos superiores a 4 segundos, Pz=0 debería disminuir. Alternativamente, si Pz=0 se debía a presión en una región elástica aguas abajo, entonces debería haber una caída progresiva de presión conforme aumenta el tiempo hasta Pz=0. Tercero, un modelo matemático, en el que se construyó un modelo para determinar si una estimación razonable de la compliance vascular podría explicar el Pz=0 observado en la segunda prueba con tiempos variables hasta flujo cero.
Materiales & Metodos
Preparación quirúrgica
- Dieciocho perros mestizos con un peso de 15-22 kg fueron estudiados
- Anestesiados con 25 mg/kg de pentobarbital sódico
- Se suplementó con 0.5-1 mg/h según fuera necesario
- Los animales fueron intubados y ventilados con un respirador de volumen (volumen corriente de 12-15 ml) durante todo el experimento
- Se suplementó oxígeno según fuera necesario
- Solo se incluyeron animales cuyo hematocrito >35%.
- Se aisló y canuló la arteria femoral derecha para monitorear Pa y extraer sangre para el análisis de gases sanguíneos
- La vena femoral fue canulada para la administración de medicación
- Se infundió lentamente solución salina al 0.9% para compensar las pérdidas de volumen durante el experimento
- La arteria femoral izquierda fue aislada en el triángulo femoral
- Se ligó y se insertó una cánula metálica con un puerto lateral para la medición de presión (Pa) en el extremo distal del vaso, más allá de la ligadura.
- La cánula se conectó a una sonda de flujo electromagnética
- Estaba unida a un conector en «Y»
- Un brazo de la «Y» se conectó a una cánula en el segmento proximal de la arteria femoral, por encima de la ligadura
- El otro se conectó a una línea que estaba unida a una bomba que extraía sangre de la arteria carótida izquierda
- Se utilizó una aguja espinal de calibre 16 para pasar hilos quirúrgicos a través del muslo y por debajo de la arteria femoral, la vena y el nervio
- Estos fueron atados firmemente para eliminar el flujo colateral hacia la pierna
- Finalmente, Qin se detuvo temporalmente, y la vena femoral fue ligada en la mitad del triángulo femoral
- El extremo distal fue canulado, y un puerto lateral del catéter se utilizó para medir la presión
- En algunos animales, se colocó otra sonda de flujo en la cánula
- Luego, el flujo venoso pasaba a través de un tubo colapsable que podía elevarse o bajarse para controlar Pv; el flujo sanguíneo se devolvía posteriormente a la vena femoral proximal.
Mediciones
- Las presiones fueron medidas con transductores Trantec, conectados a preamplificadores y a un registrador de ocho canales (modelos 8805A y 7418A).
- La señal de flujo y la presión arterial fueron registradas en un grabador de cinta de cuatro canales para su posterior análisis en un IBM-PC equipado con un convertidor analógico-digital y programado para calcular la relación presión-flujo.
Procedimientos
Mediciones dinámicas de presión-flujo:
- En ocho animales, la extremidad amputada fue perfundida con sangre proveniente de la arteria femoral proximal para que se pudiera determinar el requerimiento de flujo «natural» de la extremidad.
- Luego, se clampeó la línea proximal y se ajustó la bomba para proporcionar el mismo flujo medio que el flujo natural.
- Posteriormente, el flujo de la bomba se redujo manualmente a cero a una velocidad constante durante aproximadamente 3 segundos y se mantuvo en cero durante 6 segundos.
- Luego, el flujo se restauró al nivel de control durante aproximadamente 2 segundos y luego se redujo nuevamente a cero para evaluar la relación presión-flujo con hiperemia reactiva (Figura 1).
- Se obtuvieron cuatro o cinco mediciones para cada perro.
- A continuación, se produjeron contracciones musculares mediante voltajes supramáximos, con estímulos de 5 Hz y una duración de 15 mseg.
- Cuando el flujo se había estabilizado, se repitió el mismo procedimiento con una reducción del flujo a cero, detención del flujo durante 6 segundos y una segunda prueba «reactiva».
Presiones venosas variables:
- Estos estudios se realizaron con Qi constante o Pa constante en 10 perros.
- Qi fue proporcionado por la bomba.
- Primero, se obtuvieron tres mediciones de Pz=0 reduciendo el flujo a cero en un período de 3 segundos y restaurándolo al nivel de control.
- Luego, se elevó el resistor de Starling en la salida venosa en pasos, y se registraron los cambios en Pa.
- En algunos de estos estudios, Pz=0 también se registró reduciendo el flujo a cero a diferentes niveles de Pv.
- Para Pa constante, Pv se modificó como se describió anteriormente, y la perfusión se proporcionó desde la arteria femoral proximal nativa, que no se vio afectada por el aumento de Pv en la pierna y, por lo tanto, representa estudios con presión constante.
- En estos estudios, se registraron los cambios en Qin.
- Tanto los estudios con flujo constante (n=5) como los estudios con presión constante (n=6) con cambios en la presión venosa también se realizaron después de la inyección de 10 mg de fentolamina en la arteria femoral.
Tiempo variable hasta flujo cero:
- Se obtuvieron mediciones dinámicas de presión-flujo reduciendo el flujo a cero en un período de 1 a 10 segundos y luego restaurándolo al nivel de control a la misma velocidad en seis perros.
- Se obtuvieron entre siete y quince mediciones por cada animal, y los valores de Pz=0 se graficaron en función del tiempo hasta flujo cero.
Análisis de la relación presión-flujo dinámica
- Los datos de los estudios dinámicos de presión-flujo fueron digitalizados en un IBM-PC que muestreaba la señal a 50 Hz.
- Qin se graficó contra Pa, y se calculó una ecuación de regresión lineal.
- Hubo una pequeña fase inicial curvilínea y luego una relación lineal.
- El programa proporcionó la pendiente de la fase lineal y Pz=0, que es la intersección con el eje x de la ecuación (ver Figura 1).
- Como se puede observar en la señal analógica, la presión continuó disminuyendo cuando el flujo de entrada fue cero, pero la línea de presión-flujo incluyó solo los puntos hasta el momento en que el flujo de entrada llegó a cero.
Estadísticas
- Los datos se presentan como media ± desviación estándar (SD).
- Cuando se obtuvieron mediciones repetidas, se utilizó un análisis de varianza para mediciones repetidas para determinar primero si existía una variación significativa en la población; cuando se encontraron diferencias significativas entre grupos, estas fueron analizadas con una prueba de Newman-Keuls.
- Las relaciones entre variables lineales fueron analizadas mediante análisis de regresión lineal.
Resultados
Relaciones presión-flujo dinámicas.
- En los ocho perros en los que se estudiaron la hiperemia reactiva y las contracciones, la Pa inicial en reposo fue de 131 ± 21.6 mmHg, y con estimulación de 5 Hz fue de 123.1 ± 17.3 mmHg.
- El flujo de control fue de 139.8 ± 48.6 ml/min, y con estimulación de 5 Hz aumentó a 250.3 ± 43.5 ml/min (p < 0.01).
- La Pv de control fue de 3.9 ± 1.4 mmHg, y con estimulación de 5 Hz aumentó a 4.9 ± 2.0 mmHg.
- Cuando Qin en la extremidad amputada en reposo se redujo a cero en un período de 2-3 segundos, la línea de presión-flujo resultante fue en su mayoría lineal, aunque en algunos casos la relación fue curvilínea, con convexidad hacia el eje de presión en flujos bajos.
- Un ejemplo se muestra en Figura 1, y los datos promedio se presentan en Figura 2.
- El Pz=0 promedio fue de 71 ± 16 mmHg, y el coeficiente de correlación medio (r) fue 0.96 ± 0.03.
- Es importante señalar que Pa continuó disminuyendo después de Pz=0, y aunque esto no fue estudiado sistemáticamente, los estudios posteriores dejaron claro que el flujo venoso continúa cuando Qin es cero.
- Cuando Qin se restauró al nivel de control después de 6 segundos de oclusión y se repitió la maniobra, Pz=0 disminuyó a 64 ± 16 mmHg.
- Con contracciones de la extremidad amputada, Pz=0 disminuyó a 50 ± 18 mmHg y a 40 ± 19 mmHg en la medición reactiva después de 6 segundos de oclusión vascular.
- Los datos de los ocho animales se resumen en Figura 2.
- La pendiente de las líneas de presión-flujo incremental se volvió progresivamente más pronunciada, y Pz=0 se hizo más baja con la prueba «reactiva», las contracciones musculares y la combinación de la prueba reactiva con contracciones.
- De manera similar al enfoque de Aversano, las relaciones presión-flujo también se obtuvieron disminuyendo y aumentando el flujo de manera lineal para intentar eliminar los efectos de capacitancia, ya que Aversano propusieron que una línea presión-flujo «libre de capacitancia» se encuentra entre las fases descendente y ascendente de la relación presión-flujo.
- Además, se observó la presión a medida que el flujo se incrementó al doble del flujo de control.
- Un ejemplo con la fase descendente primero, seguida por la ascendente, se muestra en Figura 3 (panel izquierdo), y la fase ascendente primero, seguida por la descendente, se muestra en Figura 3 (panel derecho).
- La fase ascendente después de una fase descendente en la relación presión-flujo se desplazó hacia la izquierda, pero permaneció lineal, de modo que Pa fue menor que la Pa de control cuando Qin volvió a su nivel de control.
- Como se puede ver en el ejemplo, el desplazamiento dependió del tiempo en que Qin estuvo cero (es decir, mayor en el ejemplo inferior que en el superior).
- Cuando la fase ascendente fue primera, hubo un pequeño desplazamiento a la izquierda durante la fase descendente y un desplazamiento mayor a la izquierda con la siguiente fase ascendente.
Cambios en la presión venosa central.
Flujo constante::
- Las Figuras 4A y 4B muestran gráficas de Pa versus Pv en 10 perros, en los que Qin se mantuvo constante y Pv se elevó en pasos.
- Estas gráficas se presentan por separado únicamente para mayor claridad.
- En algunos perros (por ejemplo, 3, 5 y 7), Pv pudo aumentarse por encima de 20 mmHg antes de que Pa cambiara, mientras que, en otros perros, pequeños cambios en Pv produjeron aumentos en Pa.
- Los resultados promedio se muestran en la Figura 5.
- El cambio promedio en Pv antes de que ocurriera un cambio en Pa fue de 9.6 ± 6.2 mmHg; el Pz=0 para estos animales fue de 56.9 ± 11.7 mmHg (basado en una disminución de 4 segundos en Qin).
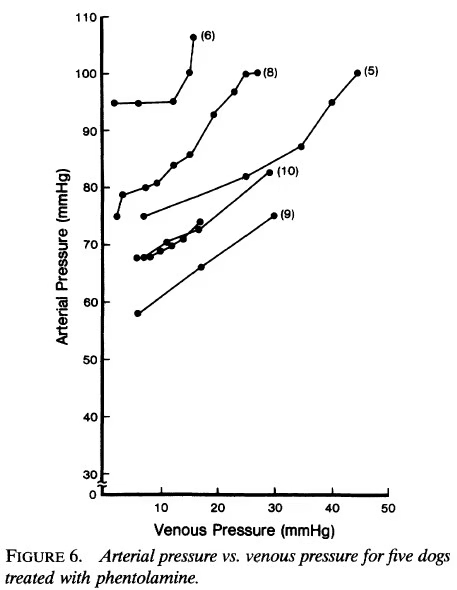
- En cinco animales tratados con fentolamina, el cambio en Pa en respuesta a un cambio en Pv fue más consistente (Figura 6), y en solo uno de estos perros (perro 6) Pv pudo elevarse más que unos pocos milímetros de mercurio sin un aumento en Pa; el promedio fue de 2.6 ± 4.3 mmHg.
- En estos cinco animales, Pz=0 fue de 35.6 ± 9.7 mmHg con fentolamina y de 54.1 ± 26.9 mmHg sin ella.
Presión constante:
- Los cambios en Qin con variaciones en Pv se muestran en la Figura 7A.
- En la mayoría de los animales, Pv pudo incrementarse ligeramente antes de que Qin disminuyera.
- El cambio promedio (Figura 5) en Pv antes de que ocurriera un cambio en Qin fue de 6.8 ± 7.3 mmHg, con un rango de 0-20 mmHg; la Pv promedio en el punto de cambio fue de 14.0 ± 8.1 mmHg.
- Después del tratamiento con fentolamina (Figura 7B, n=6), el cambio en Pv antes de que Qin disminuyera fue de 2.2 ± 3.9 mmHg.
- Pz=0 aumentó cuando Pv se incrementó.
- Un ejemplo se muestra en la Figura 8.
- En cuatro perros, con un aumento promedio en Pv de 28.7 ± 4.5 mmHg, Pz=0 aumentó en 17.6 ± 10.1 mmHg.
- En cada animal, la pendiente de la relación presión-flujo disminuyó, lo que indica un aumento en la resistencia.
- En tres perros, Pv se elevó hasta que Qin se detuvo.
- En cada caso, Qin se volvió cero cuando Pv = Pa, a pesar de que Pz=0 aumentó con el incremento de Pv.
Variable de tiempo a flujo cero
- Los efectos de la variación del tiempo hasta Pz=0 en seis animales se muestran en la Figura 9.
- Hubo una caída inicial marcada en Pz=0 con tiempos crecientes hasta flujo cero, seguida de un cambio mínimo entre 5 y 10 segundos.
- Los valores iniciales de Qin, Pa y Pv de cada animal se utilizaron para calcular la compliance predicha con modelos de uno y dos compartimentos.
- El modelo de un solo compartimento incluyó una resistencia, con compliance y resistencia en serie (Figura 9A).
- La disminución en Qin fue lineal, según el protocolo.
- La solución de esta ecuación dio:

- donde β es la tasa de cambio del flujo de entrada, Rout es la resistencia de salida, C es la capacitancia, Pb es la presión de retorno (presión aguas abajo) y Qin (0) es el flujo inicial.
- Los valores experimentales de Qin (0), Pz=0, Pv y β se utilizaron para calcular la compliance, suponiendo diferentes proporciones de resistencia proximal versus distal.
- Primero, se usaron los datos del perro en la Figura 10E para evaluar el modelo.
- Como era de esperarse en un modelo con una sola resistencia y compliance, cuando Pv se usó como Pb, la solución requirió un Rout alta o una C grande para explicar un Pz=0 de 35 mmHg cuando el tiempo para alcanzar flujo cero fue de 10 segundos.
- Con Rout representando el 30% de la resistencia total, la compliance debía ser 4 ml·mmHg⁻¹, lo que requería un volumen estresado de 150 ml desde la arteria femoral de una sola extremidad posterior hasta la presión de salida venosa.
- No se encontró una solución cuando Rout fue menor al 30% de la resistencia total.
- Con el 100% de la resistencia distal a la región de compliance, C se estimó en 0.4 ml·mmHg⁻¹, lo que dio un volumen de 5 ml para la vasculatura de la extremidad posterior.
- Cuando este valor de C se usó para calcular Pz=0 para tiempos hasta flujo cero menores a 10 segundos, la curva fue mucho más pronunciada de lo observado experimentalmente.
- También se probó un modelo de dos compartimentos (Figura 9B). En este modelo, el sistema comienza con una pequeña resistencia arterial que vacía en una región de baja compliance (arterias), que luego vacía en una región de alta resistencia (arteriolas), y posteriormente en una región de alta compliance (vénulas y venas).
- La segunda compliance se varió entre 0.2 y 0.8 ml·mmHg⁻¹·kg⁻¹, y la primera compliance se varió entre 1/10 y 1/30 de la segunda compliance.
- Las resistencias se distribuyeron como R₁ = 5%, R₂ = 80% y R₃ = 15% de la resistencia total.
- Una vez más, los valores experimentales de Qin y Pv fueron utilizados, y la serie de ecuaciones que describen este modelo se analizaron por reiteración en una computadora PC, con tiempos hasta flujo cero entre 1 y 10 segundos, como se obtuvo experimentalmente.
- Sin embargo, este modelo tampoco logró predecir los resultados experimentales (Figura 10).
- Finalmente, en el modelo de un solo compartimento, el valor de Pb en la Ecuación 1 se fijó en un nivel ligeramente inferior al Pz=0 medido a 10 segundos.
- La justificación para esto fue que este valor podría representar una presión de salida crítica a nivel de las arteriolas (Figura 9C).
- El valor medido de Pz=0 a 2 segundos se usó en la Ecuación 1 para calcular C. Esta C y la ecuación se usaron para predecir otros valores de Pz=0 con diferentes velocidades de reducción de flujo.
- Los parámetros utilizados para los cálculos en los seis perros de la Figura 11 se muestran en Tabla 1.
- Para estos cálculos, el 100% de la resistencia se situó distal a la región de compliance, lo que en este caso representaría la compliance arterial.
- Esto se justificó porque 1) la presión crítica es tan alta que la compliance debe estar en las arterias y hay poca resistencia significativa en las arterias grandes, y 2) la relación entre Pz=0 y el tiempo hasta flujo cero parecía extrapolarse hasta la presión arterial.
- Cuando la resistencia se redujo al 95% del total, los resultados apenas mostraron diferencias.
- Como se muestra en la Figura 11, este modelo predijo Pz=0 de manera casi perfecta.
- Esta figura también muestra los valores de presión arterial cuando el flujo se detuvo por diferentes períodos de tiempo.
- Estos valores cayeron considerablemente por debajo de los valores obtenidos con la reducción gradual del flujo.
Discusión
Las principales observaciones de este estudio pueden resumirse de la siguiente manera:
- Cuando el flujo se redujo rápidamente a cero (es decir, en 1-3 segundos), la presión en flujo cero (Pz=0) fue mucho mayor que Pv.
- Pz=0 disminuyó con la hiperemia reactiva, el ejercicio y el tratamiento con fentolamina.
- Pz=0 disminuyó considerablemente cuando el tiempo hasta flujo cero aumentó de 1 a 4 segundos, pero mostró poca variación a partir de los 10 segundos.
- Detener repentinamente el flujo resultó en un Pz=0 mucho menor que cuando el flujo se redujo gradualmente.
- Las presiones venosas pudieron aumentarse aproximadamente 10 mmHg, hasta 14-16 mmHg, antes de que afectaran el flujo de entrada con presión arterial constante o la presión arterial con flujo constante.
Se deben considerar varios factores técnicos. Es fundamental evitar el flujo colateral (Hoffman, 1985), ya que las colaterales arteriales podrían mantener la presión en flujo cero, mientras que las colaterales venosas reducirían los efectos del aumento de la presión venosa. Para eliminar estos flujos colaterales, se colocó un torniquete ajustado alrededor de la pierna; su efectividad se verificó observando qué tan baja caía Pa después de un período de oclusión arterial, así como midiendo cualquier efluente de la línea de presión abierta con la oclusión del flujo de entrada. Solo se incluyeron preparaciones con flujo colateral mínimo (es decir, <5% del valor basal). Sin embargo, estas pruebas no pudieron repetirse a lo largo del experimento, ya que afectaban la preparación, por lo que el flujo colateral podría haber aumentado en algunos animales durante el experimento. Además, los reflejos y el tono vascular se mantuvieron intactos, lo que significa que los mecanismos miogénicos o los reflejos venoarteriales (Baez, 1974; Baez, 1977; Honig, 1979; Henriksen, 1983; Phillips, 1955) podrían haber influido en Pa o Qin durante los cambios en Pv. No obstante, fue necesario estudiar los animales con tono y reflejos intactos, ya que se cree que las presiones críticas dependen del tono arteriolar (Klocke, 1981; Klocke, 1985).
Como en estudios previos sobre las relaciones dinámicas presión-flujo, Pa seguía siendo alta cuando el flujo arterial de entrada alcanzaba cero. En los estudios iniciales con tiempos hasta flujo cero de 2-3 segundos, Pz=0 fue de 71 ± 16 mmHg. Este valor es mucho mayor al observado por Ehrlich en la extremidad amputada del perro, pero en su estudio primero obstruyeron el flujo por más de 30 segundos para inducir hiperemia reactiva. Después de 6 segundos de detención del flujo, Pz=0 fue menor en este estudio y disminuyó aún más cuando se estimularon contracciones musculares. La pregunta sigue siendo si esto puede explicarse mediante un mecanismo de resistencia de Starling o si se debe simplemente a un efecto de capacitancia.
Tiempo a flujo cero
Con tiempos hasta flujo cero de 1-4 segundos, hubo una disminución exponencial en Pz=0, seguida de pocos cambios en Pz=0 hasta 10 segundos. El primer modelo utilizado para analizar estos resultados incluyó una resistencia, seguida de una región con compliance, que se vaciaba a través de otra resistencia (Figura 9). Cuando Pv se utilizó como la presión de salida (Pb), la compliance calculada fue demasiado alta en comparación con lo que se esperaría en la extremidad (Diana, 1973). Por otro lado, solo se pudo obtener una estimación más razonable de compliance si la mayor parte de la resistencia se encontraba distal a la región complaciente. Esto requeriría que la región con compliance estuviera en las arterias en lugar de las venas, pero en ese caso, la compliance calculada resultó demasiado grande para una compliance arterial. Finalmente, cuando el valor de compliance obtenido con la rampa de 10 segundos se introdujo en la ecuación para tratar de predecir Pz=0 en otros tiempos hasta flujo cero, Pz=0 fue sobreestimada en tiempos cortos y subestimada en tiempos largos. Más importante aún, no hubo ningún valor de compliance que pudiera haber predicho el estancamiento de Pz=0 con tiempos crecientes hasta flujo cero, tal como se observó (Figuras 10 y 11). El modelo no mejoró al agregar una segunda compliance en serie con la primera (Figura 10). Sin embargo, los datos fueron descritos casi perfectamente utilizando un Pb que era mucho mayor que Pv y muy cercano a Pz=0 a los 10 segundos, tal como ocurriría con un mecanismo de resistencia de Starling. En este caso, la pendiente de la línea presión-flujo está determinada por la resistencia entre Pa y esta presión de salida, a la que llamaremos presión crítica (Pb), para distinguirla de Pz=0, que es la presión observada en flujo cero. Distal a esta Pb, o resistencia de Starling, se encuentran los capilares, la capacitancia y la resistencia venosas.
Es importante señalar que, mientras la presión capilar sea menor que Pb, la resistencia venosa no contribuirá a la resistencia al flujo de entrada, y el flujo estará simplemente determinado por la ecuación:

Un problema con este análisis surge cuando se examinan los experimentos en los que el flujo de entrada se detuvo abruptamente, ya que Pz=0 cayó a niveles mucho menores que el Pb determinado por las reducciones graduales de flujo de hasta 10 segundos (Figura 11). La hipótesis del resistor de Starling predice que, incluso con una detención repentina del flujo, Pa solo debería caer hasta Pb. Sin embargo, esta observación puede explicarse fácilmente por la presencia de colaterales arteriales, que pueden eludir Pb, requiriendo una conductancia de solo 1.5% del total. Por lo tanto, cuando Pa cae al nivel de Pb, el flujo puede continuar a través de estas colaterales, pero debido a la baja conductancia, la presión disminuye muy lentamente. Este valor de conductancia se puede utilizar para predecir que, como se observa, Pz=0 a los 30 segundos sigue siendo mayor que Pv. Cabe destacar que la conductancia que determina la caída en Pa es la conductancia a través de las colaterales, que es solo 1.5% del total. Los datos de los estudios con tiempos variables hasta flujo cero pueden explicarse muy bien mediante un mecanismo de resistencia de Starling, junto con la coexistencia de una pequeña cantidad de flujo colateral. Estos resultados también muestran que la compliance arterial tiene un efecto significativo en Pz=0 cuando los tiempos hasta flujo cero son cortos (<4 segundos).
Cambios en la presión venosa central
Aunque los datos sobre los tiempos variables hasta flujo cero pueden explicarse bien mediante un mecanismo de resistencia de Starling, los resultados de los cambios en Pv con flujo constante o Pa constante podrían parecer, en un primer análisis, contradictorios con esta hipótesis. Esto se debe a que Pv solo pudo aumentarse en aproximadamente 10 mmHg (hasta 14-16 mmHg) antes de que Pa o el flujo aguas arriba se vieran afectados, a pesar de que Pz=0 se encontraba entre 50 y 60 mmHg. Parte de esta discrepancia puede explicarse por los efectos de capacitancia mencionados anteriormente, ya que cuando se tiene esto en cuenta, Pb resultó ser de 42 mmHg en comparación con 54 mmHg (Tabla 1) en los seis animales con tiempos variables hasta Pz=0. Otro factor es que Pb representa el valor medio de una familia de presiones críticas (Mitzner, 1974), por lo que los cambios de presión en la región aguas abajo podrían transmitirse a través de canales con presiones críticas bajas. Sin embargo, si esta fuera la única explicación, la variabilidad de la presión en la región de compliance en cada lecho vascular individual habría sido muy grande. Existe, sin embargo, una explicación más simple que sigue siendo completamente consistente con la hipótesis de resistencia de Starling y que coincide exactamente con lo predicho en el artículo clásico de Permutt y Riley sobre la «cascada vascular». Debe recordarse que la parte inferior de la cascada, o resistor de Starling, puede ser una presión antes de los capilares y no necesariamente la presión en las venas. Dado que hay una resistencia entre estos dos puntos, esta presión puede ser mayor que Pv, y es la diferencia entre Pb y esta presión la que determina la altura de la cascada. Si la diferencia de presión de la cascada vascular fue de aproximadamente 10 mmHg, como lo sugiere el cambio promedio en Pv antes de que hubiera un efecto aguas arriba, y Pb fue 42 mmHg, entonces esta presión sería de aproximadamente 32 mmHg, lo que está dentro del rango de presiones observadas en la región precapilar y es similar al valor predicho por Permutt y Riley en su análisis de los datos de Burton.
Otro resultado que necesita explicación es el aumento de Pz=0 con el aumento de Pv. Bellamy et al. explicaron esto a través de un aumento en la presión tisular, que podría comprimir la región de presión crítica e incrementar Pb. Ahora, este resultado puede explicarse más fácilmente al notar que un aumento suficiente en Pv puede elevar la presión debajo de la cascada vascular hasta Pb, lo que elimina el efecto de cascada y comienza a afectar las características vasculares aguas arriba (Figuras 4-7). Dado que el flujo se mantuvo constante con una bomba, cuando el aumento de Pv superó la cascada, hubo un aumento en Pa. Pz=0 también aumentó, ya que está influenciado por Pa. El segundo punto importante proviene nuevamente de Permutt y Riley. Cuando Pv se eleva por encima de la cascada, la resistencia venosa debe sumarse a la resistencia arterial, lo que significa que la resistencia total aumenta y la pendiente de la línea presión-flujo debe disminuir, lo cual es exactamente lo que se observó en Figura 8 en todos los casos. Finalmente, Pz=0 disminuyó con hiperemia reactiva y fentolamina. Hay dos factores que podrían explicar esto. Primero, la caída en Pa que ocurrió con ambas maniobras habría provocado una disminución en Pz=0. Segundo, probablemente hubo una reducción del tono arteriolar, lo que disminuyó la cascada vascular. De hecho, después del tratamiento con fentolamina, el aumento en Pv tuvo un efecto inmediato sobre el flujo o la presión aguas arriba en todos los perros, excepto uno. La pequeña reducción en Pa durante la contracción muscular también pudo haber contribuido a la disminución de Pz=0 en la extremidad posterior en ejercicio, así como una reducción de la presión en la región de compliance.
Dole et al. propusieron una explicación alternativa para Pz=0. Sugirieron la presencia de una resistencia media variable, que aumentaría progresivamente a medida que el flujo disminuye. Esto podría deberse al colapso de los vasos y a una pérdida del área transversal, como lo plantea Burton en su concepto de presiones críticas de cierre. Sin embargo, Dole et al. no explicaron por qué la resistencia disminuye a una tasa que produce una disminución lineal en la relación presión-flujo. Además, aunque su análisis predice correctamente el desplazamiento hacia la derecha de la relación presión-flujo con el aumento de Pv, no predice la disminución en la pendiente que se observó en este estudio. Finalmente, tampoco explica de manera clara la diferencia entre la disminución gradual del flujo y la oclusión repentina del flujo. Otras explicaciones que se han propuesto para explicar las presiones críticas vasculares incluyen efectos del hematocrito (Whittaker, 1933), tensiones interfaciales (Sherman, 1981), mecanismos miogénicos (Hanson, 1962; Smiesko, 1971; Johnson, 1976) y ajustes reflejos (Baez, 1974; Baez, 1977; Honig, 1979; Henriksen, 1983).
Significado del mecanismo del resistor de Starling
Aunque estos datos respaldan la presencia de un mecanismo de resistencia de Starling en los vasos arteriales, el efecto de capacitancia observado también tiene implicaciones importantes en la evaluación de Pz=0. Por lo tanto, cualquier medición de presión-flujo con menos de 4 segundos para la estabilización tendrá un componente significativo de capacitancia, lo que significa que las mediciones de Pz=0 realizadas por Dewey et al. y Early et al. en la circulación cerebral, Ehrlich et al. en el riñón y la extremidad amputada (Ehrlich, 1980), y Bellamy en la circulación coronaria sobrestimarían significativamente Pb. Se pueden extraer las mismas conclusiones sobre los estudios de autorregulación, como los de Mosher et al. en la circulación coronaria, en los cuales algunos de los efectos autorreguladores podrían haber sido en realidad efectos de capacitancia. Además, las mediciones denominadas «libres de capacitancia» de Aversano et al., Canty et al. y Dole y Bishop podrían no haber estado completamente libres de capacitancia, ya que utilizamos una técnica similar a la de Aversano et al. y encontramos que Pz=0 seguía siendo mayor que el Pb predicho para tiempos hasta flujo cero de 5-10 segundos.
La presencia de un mecanismo de resistencia de Starling tiene importantes implicaciones en la comprensión de la circulación periférica. Primero, implica que el flujo hacia diferentes regiones está determinado por Pa – Pb y la resistencia entre ambos, en lugar de Pa – Pv y la resistencia total. Esto significa que la resistencia efectiva es menor que la resistencia total, y que el ajuste de Pb es un factor clave en el control del flujo a diferentes regiones del cuerpo. Es decir, el flujo puede modificarse tanto por cambios en Pb como por cambios en la resistencia arterial. La presencia de un mecanismo de resistencia de Starling también implica que la presión capilar, y por lo tanto las presiones de filtración, pueden ajustarse sin afectar el flujo de entrada a la región, ya que las presiones capilares están por debajo de la cascada vascular. Esto sugiere que el uso de la relación entre resistencias aguas arriba y aguas abajo para evaluar la filtración capilar no es en realidad un método preciso para estimar esta presión. La existencia de una cascada vascular a nivel arteriolar también implica que los cambios en Pv provocados por maniobras respiratorias como la inspiración, la maniobra de Valsalva o la tos no necesariamente afectarán el flujo al lecho capilar y, por lo tanto, este mecanismo contribuye a mantener la estabilidad del flujo sanguíneo. Finalmente, la presencia de una presión crítica significa que, ante una disminución repentina del gasto cardíaco, Pa no disminuirá tan rápidamente como lo predicen los análisis que no incluyen un mecanismo de resistencia de Starling. Esto ayudará, al menos temporalmente, a mantener el flujo en áreas críticas como el cerebro, que probablemente tiene una presión crítica más baja que el músculo en reposo.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}